Cet article en deux volets présente la bioélectrochimie ainsi que son application à l'étude de phénomènes cellulaires.
Le premier volet présente la bioélectrochimie et ses outils essentiels pour sonder le vivant.
Le deuxième volet (ci-dessous) présente deux exemples de l'utilisation de ces outils à l'étude des phénomènes cellulaires.
Premier exemple : effet de la courbure membranaire
Facteurs physico-chimiques
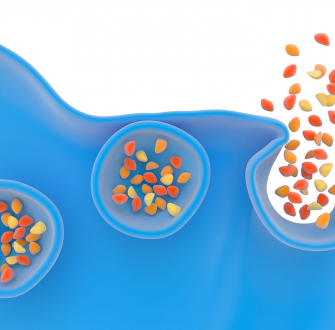
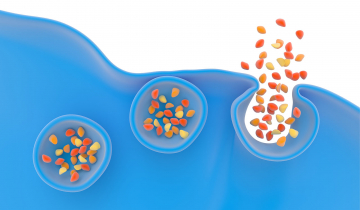
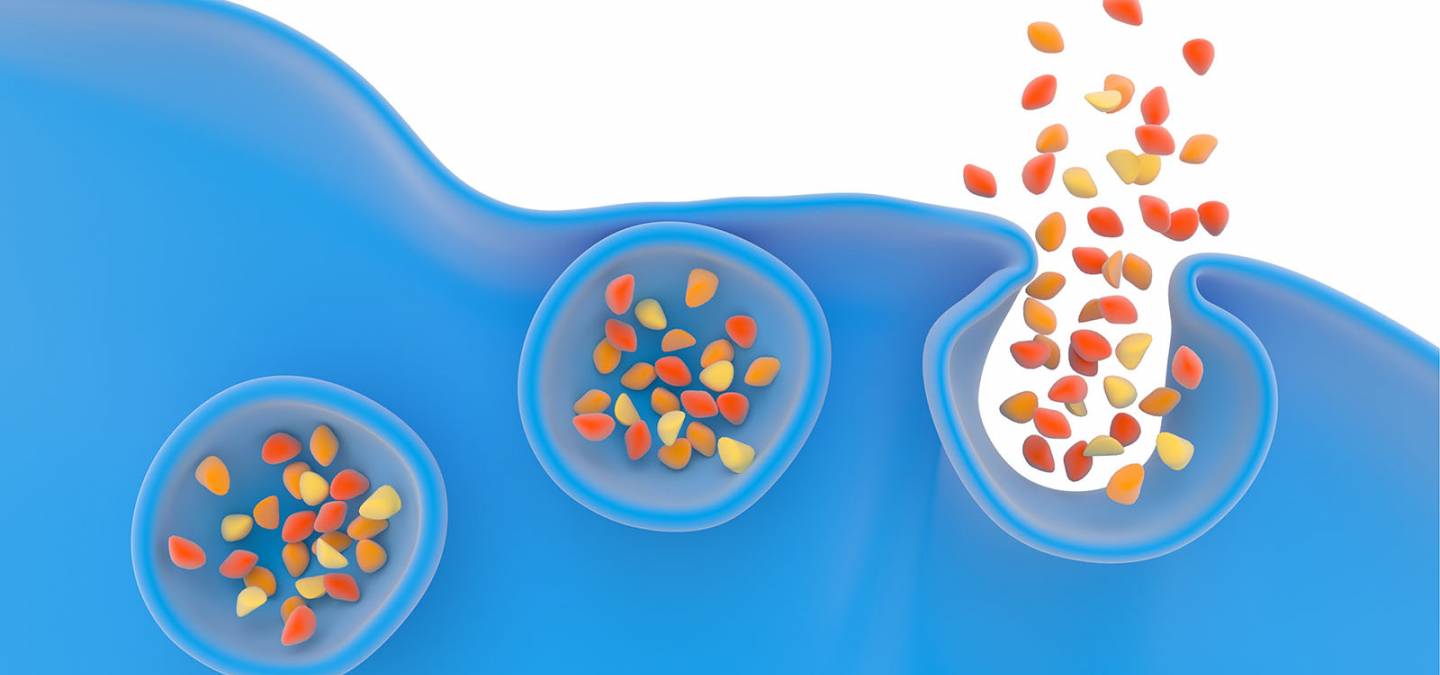
Des expériences ont été menées pour étudier le rôle de la courbure membranaire dans la formation du pore de fusion et son influence sur la sécrétion cellulaire, la vitesse de libération des biomolécules ainsi que la quantité d'espèces relarguées. Ces expérimentations ont été réalisées selon la configuration expérimentale décrite ci-dessus et sur cellules vivantes, des cellules chromaffines issues de bovins. Ces cellules chromaffines ont été baignées dans des solutions de phospholipides (le constituant lipidique important des cellules animales et végétales, comportant une partie polaire du côté du milieu aqueux – extracellulaire ou intracellulaire – et une partie grasse ou hydrophobe au cœur de la membrane). Ces phospholipides ou acides gras ont été choisis avec une géométrie particulière afin qu'ils modifient la courbure membranaire lors de la formation du pore de fusion et sont incubés avec les cellules une durée donnée afin qu'ils s’insèrent dans la membrane cellulaire externe. L’originalité de ces essais réside dans le fait que des travaux ont déjà montré en 1995 le rôle de la courbure membranaire sur la fusion membranaire mais uniquement sur des membranes lipidiques modèles synthétiques et non pas sur des cellules vivantes réelles [1].
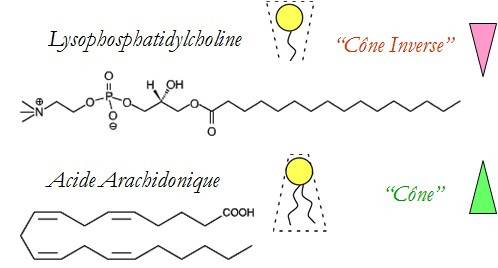
LPC avec une grosse tête polaire et une chaîne hydrophobe
Les géométries des molécules choisies sont de deux types : l’une s’appelle la lysophosphatidylcholine, notée LPC, et l’autre est un acide gras, l’acide arachidonique, noté AA. La LPC a une forme de « cône inverse » qui contraint la courbure membranaire et l’accentue, favorisant ainsi la fusion membranaire et donc la fusion des vésicules de sécrétion avec la membrane cellulaire (Figure 1). On dit alors que le LPC est fusogène. À l’inverse, l’AA a une géométrie conique qui empêche la fusion membranaire et est dit antifusogène (Figure 2).
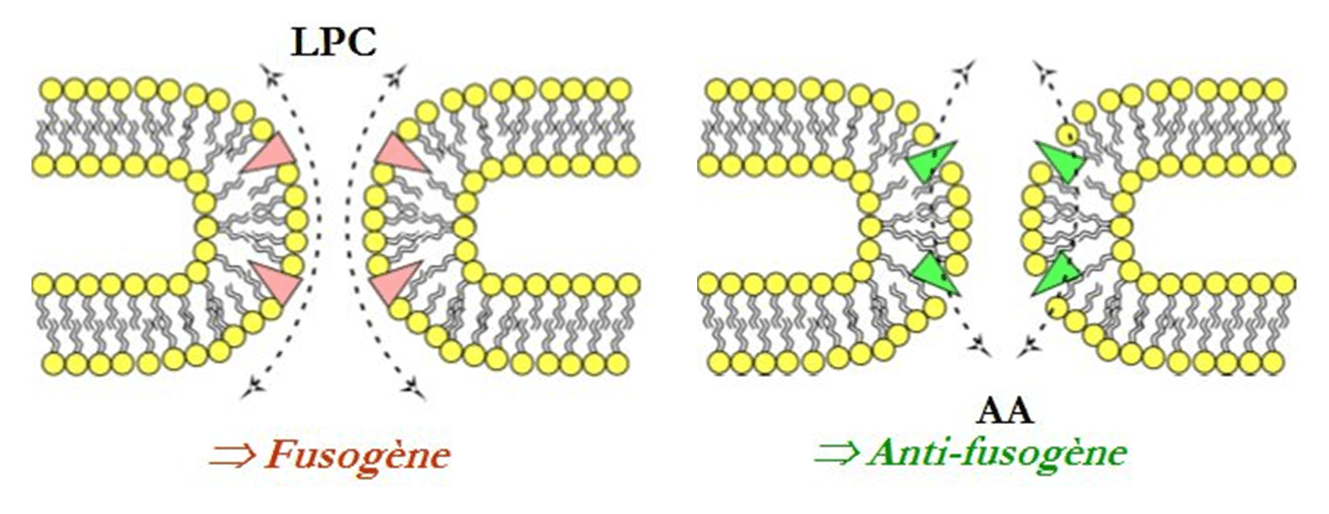
Insertion de LPC, à gauche, ou AA, à droite, dans le pore de fusion lors de l'exocytose, favorisant (effet fusogène) ou non (effet antifusogène) la fusion des membranes.
Effets sur la fréquence, la dynamique et la quantité de biomolécules de la sécrétion cellulaire
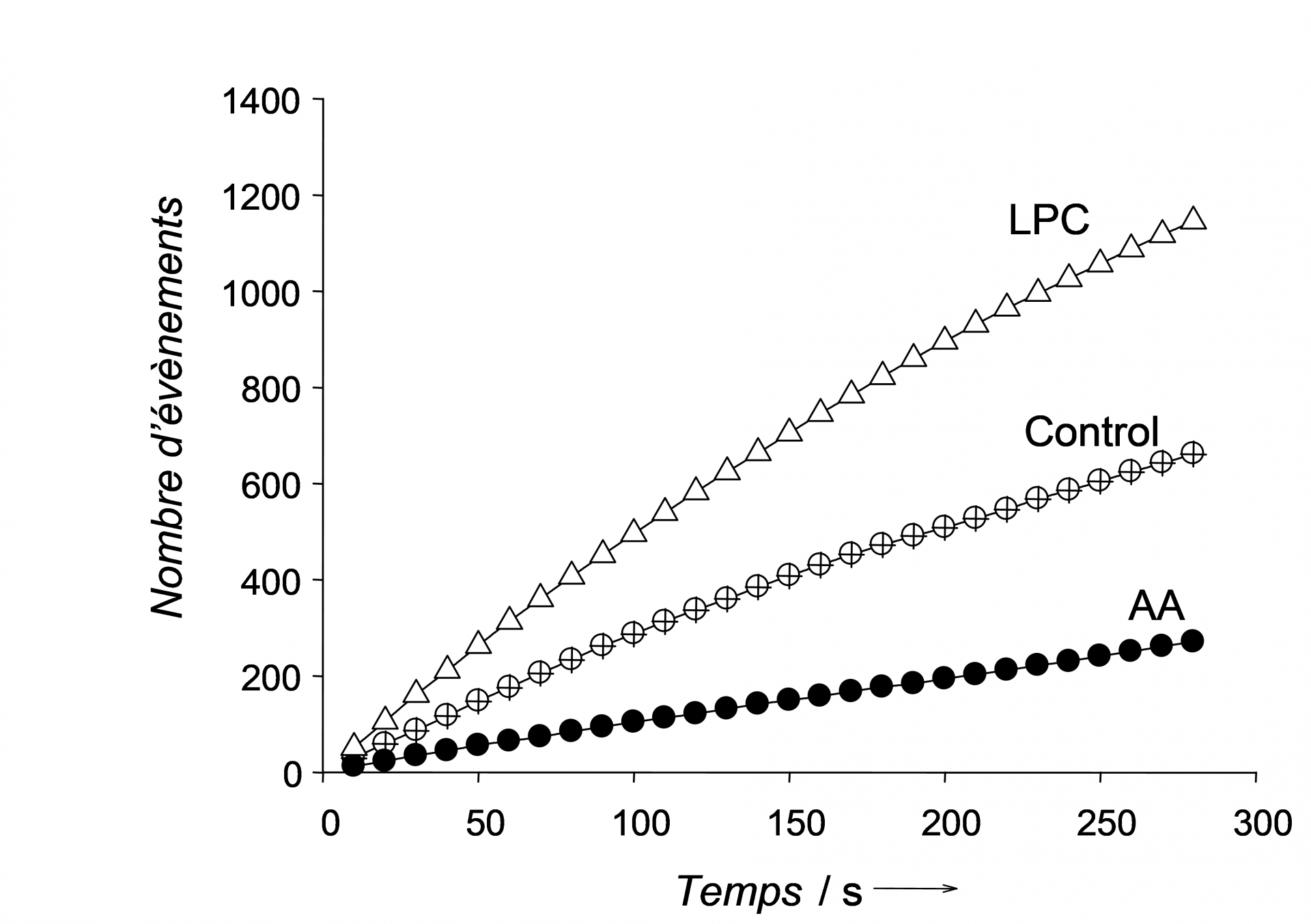
Ces expériences de modification de la courbure membranaire lors de l’exocytose ont montré que la fréquence de sécrétion est grandement affectée par la présence d’une molécule fusogène (le LPC) ou antifusogène (l’AA) dans la membrane cellulaire (Figure 3). Le nombre d’évènements détectés sur les chronoampérogrammes est en effet quasiment doublé en présence d’une molécule fusogène et divisé par un facteur d’ordre 2 en présence d’une molécule antifusogène. La vitesse de chaque événement d’exocytose a cru ainsi que le nombre de molécules libérées qui a également augmenté d’un facteur 2 en présence de la molécule fusogène. La tendance opposée est observée pour l’acide gras antifusogène.
Ces expériences démontrent sans ambiguïté que la composition membranaire lipidique et la courbure membranaire, probablement au niveau du pore de fusion, ont un rôle crucial lors de la libération de biomolécules dans l’exocytose de cellules chromaffines qui régulent la sécrétion de l’adrénaline dans le sang. Ainsi il semblerait que la présence de LPC ou de AA favorise ou défavorise l’ouverture ou la fermeture du pore de fusion créant de nouveaux événements ou en empêchant certains par rapport au contrôle. Ces résultats sont très intéressants car ils ouvrent des perspectives de traitement ou d’analyse de la neurotransmission en fonction des lipides ingérés. Par ailleurs, ils montrent que l’étude par ultramicroélectrode sur cellules vivantes dans une boite de Petri permet d’analyser très finement les facteurs physico-chimiques de contrôle de la sécrétion et d’en déterminer les effets de manière quantitative en termes de fréquence, vitesse (précision au dixième de millisecondes) et quantité de biomolécules libérées par événement1. C’est un exemple parlant de l’utilité et la puissance de ces objets micrométriques pour sonder le vivant.
Deuxième exemple : couplage d’une technique optique et d’une technique électrochimique
Principe analytique de détection optique
Les communautés de neurophysiologistes qui étudient la neurotransmission lors de l’exocytose ont une technique optique de prédilection qui n’est plus électrique, comme l’ampérométrie sur ultramicroélectrodes, mais optique. C’est une technique fluorescente qui a une résolution spatiale intéressante, de l’ordre de la centaine de nanomètres, mais une résolution temporelle médiocre par comparaison à l'ampérométrie, de l’ordre de la dizaine de millisecondes par rapport à la dynamique de l'exocytose. Cette résolution temporelle est en fait à comparer à celle de la technique électrochimique décrite dans les parties 1.5, 2 et 3 qui possède, elle, une excellente résolution temporelle (de l’ordre de la dizaine de microsecondes) par rapport à l’échelle de temps de l’exocytose, mais qui est aveugle à la détection intracellulaire et aux étapes précédant le pore de fusion.
Cette technique optique s’appelle la microscopie à onde évanescente ou microscopie de fluorescence par réflexion totale interne (dite TIRF – Total Internal Reflexion Fluorescence Microscopy – en anglais). Le principe analytique de détection de l’exocytose par TIRF est le suivant. Les vésicules de sécrétion de la cellule sont marquées au préalable par une molécule fluorescente, située dans la membrane vésiculaire ou au cœur de la vésicule, au choix. Les cellules sont déposées sur une lame de microscope fine et transparente pour permettre l’excitation optique par un laser ou une diode. Le faisceau lumineux vient frapper l’interface lame de microscope/cellule avec un angle particulier, dit angle critique. S’ensuit alors la création d’une onde évanescente de faible profondeur de pénétration, d’intensité proportionnelle à une exponentielle décroissante en fonction de la distance z dans la direction perpendiculaire au substrat (représenté par la lame du microscope - Figure 4). L’excitation de la fluorescence des vésicules se fait ensuite par cette onde évanescente qui a une profondeur de pénétration de 50 à 300 nm, quand la cellule fait plutôt une épaisseur de quelques micromètres. En conséquence, seule l’évolution de la fluorescence des vésicules proches de la membrane cellulaire adhérée est suivie, ce qui limite la pollution des signaux par les vésicules marquées loin de l’interface cellule-verre. Les évènements d’exocytose sont vus sous la forme de flash ou d’extinction de fluorescence. Les avantages de cette technique sont la détection en temps réel des vésicules, l’observation de leur mouvement avec une bonne résolution dans le plan x-y parallèle au substrat et la localisation des évènements de fusion sur la membrane cellulaire.
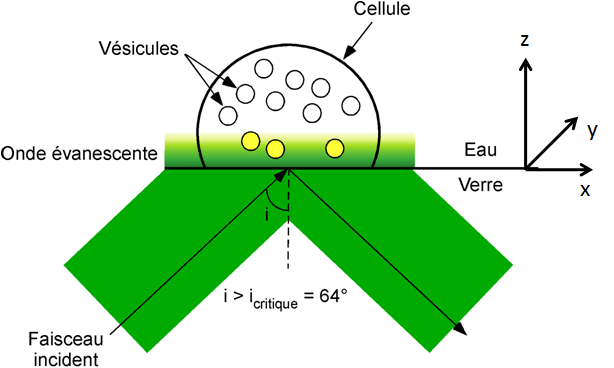
Avec l'aimable autorisation de Mr Frédéric Lemaître.
Intérêts du couplage électrochimie/fluorescence
Une fois compris les grands intérêts de chaque technique décrite plus haut : l’ampérométrie sur microélectrodes et la microscopie par onde évanescente pour le suivi de l’exocytose, il est aisé de se rendre compte de la complémentarité des deux outils analytiques et des avantages de coupler ces deux techniques en temps réel. Le couplage donnerait accès au suivi de la libération en temps réel du même évènement d’exocytose avant/pendant/après la fusion membranaire. Avant et pendant : grâce au TIRF, pendant et après : grâce aux microélectrodes. Des données quantitatives (cinétique, quantité de neurotransmetteurs) seraient obtenues avec une excellente résolution temporelle. Une cartographie des sites de libération pourrait être envisagée avec une résolution spatiale fine. Et des corrélations entre paramètres physico-chimiques et biologiques seraient tracées. Les contraintes sont néanmoins nombreuses. Il s’agit de suivre le même événement d’exocytose au même endroit de la cellule au cours du temps. Il faut donc choisir un matériau optiquement transparent et électriquement conducteur. Il faut également détecter les deux signaux à l’endroit de la cellule où celle-ci adhère sur le substrat transparent. Enfin, il faut que la sécrétion ait lieu à une fréquence modérée afin de corréler aisément les signaux des deux natures.
Détection combinée
Une première preuve de concept de couplage électrique et optique a été obtenue en 2011 avec une détection fluorescente grâce à une molécule de GFP (Green Fluorescent Protein) et une détection électrochimique de la sérotonine. Lors de cette première réalisation, le couplage a donc été réalisé avec deux sondes différentes. Le prototype a été réalisé sur lame fine et transparente d’ITO (oxyde d’indium dopé à l’étain) de 150 nm d’épaisseur. L’ITO possède des propriétés optiques, avec une bonne transmission à 80% environ dans le spectre visible, et électriques, avec une bonne conductivité électrique, et une possibilité de détecter électrochimiquement des médiateurs biologiques. Des essais ont également été réalisés sur d’autres matériaux (qui présentent une meilleure détection électrochimique des médiateurs biologiques mais qui ont une plus mauvaise transmission que l'ITO) [2]. Dans ces premières études, les deux sondes nécessaires au couplage sont différentes et en proportions relative et absolue non connues et non contrôlées, ce qui est suffisamment peu satisfaisant pour le chimiste analyticien et pour répondre aux questions posées.
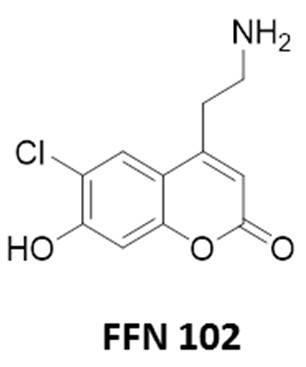
De nouveaux essais ont donc été menés avec l’utilisation de molécules nommées « faux neurotransmetteurs fluorescents » ou FNF qui ont des structures chimiques proches de neurotransmetteurs biologiques endogènes. Elles ont été initialement introduites dans la littérature spécialisée pour permettre une bonne visualisation de la neurotransmission grâce à leurs propriétés de fluorescence [3]. Certaines de ces molécules possèdent un bras amine primaire qui facilite leur internalisation dans les vésicules de sécrétion grâce à un transporteur de monoamines primaires qui se trouve dans la membrane vésiculaire. Par ailleurs, un FNF a montré des propriétés électrochimiques particulières puisqu’il est détecté électrochimiquement sur différents matériaux d’électrode comme le carbone ou l’ITO. Cette molécule, nommée FFN102, est donc une molécule de choix puisqu’elle regroupe en une sonde unique la dualité fluorescence/électroactivité tout en étant reconnue par les vésicules de sécrétion d’un point de vue biologique (Figure 5). Cette sonde duale FFN102 a permis une première étude couplée de l’exocytose sur cellules (Figure 6) et a révélé différents types de sécrétion non observés jusqu’alors [4].
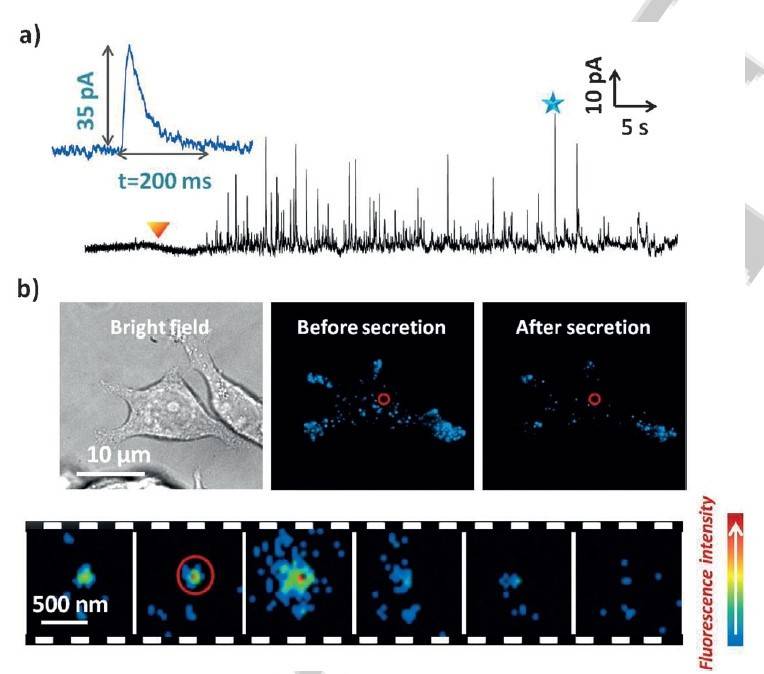
Détection combinée électrochimique a) et optique b) de la sonde duale FFN102 dans des cellules sécrétrices (exocytose détectée simultanément par a) ampérométrie et b) TIRF).
Conclusion
L'électrochimie connaît depuis quelques années un renouveau spectaculaire notamment dans le domaine des nanosciences, la bioanalyse (comme les études électrochimiques sur cellules vivantes) et la détection de courants très petits et rapides. Ces thématiques sont au cœur de cet article qui traite de la bioélectrochimie pour sonder le vivant. Pour détecter un courant, l’électrochimiste analyticien a besoin d’une électrode qui, si elle a une taille micrométrique et si elle est utilisée dans des conditions particulières, possède des propriétés intéressantes pour la détection de biomolécules au niveau de cellules vivantes uniques. Ces ultramicroélectrodes peuvent servir à analyser la sécrétion cellulaire via le phénomène d'exocytose. L’exocytose vésiculaire constitue une voie physiologique fondamentale de la communication cellulaire, utilisée par les organismes cellulaires afin de libérer hormones ou neurotransmetteurs au sein de leur environnement pour leur permettre de communiquer. L’exemple de l’influence de la composition et la courbure cellulaire membranaire est cité ici pour son grand rôle dans l’efficacité de la sécrétion.
Par ailleurs, depuis plusieurs années, les méthodes analytiques fluorescentes et électrochimiques représentent des outils de choix pour l’analyse de phénomènes biologiques. « Leur combinaison apparaît donc comme complémentaire et prometteuse pour sonder différentes facettes d’un même objet/phénomène avec des résolutions spatiale et temporelle optimisées. La notion de couplage proprement dit reste toutefois à définir et ce dernier présentera un intérêt par rapport aux deux techniques prises séparément si les informations sont recueillies simultanément et sur le même endroit de l’objet biologique. La sécrétion cellulaire illustre l’intérêt et la difficulté d’élaborer un couplage électrochimie-fluorescence sur cellule unique.» [5] Ainsi, l’ampérométrie sur électrode transparente « d’Oxyde Indium Etain » (ITO), couplée à la Microscopie de Fluorescence à Réflexion Totale Interne (TIRF), permet une analyse exhaustive (à haute résolution temporelle et spatiale) du phénomène de sécrétion nommé exocytose. Le couplage TIRF-ampérométrie a pour but de connaître, pour un même évènement d’exocytose, sa localisation et ses caractéristiques dynamiques en temps réel, ce qui implique d’adapter le dispositif expérimental. Dans ce contexte, de nouvelles sondes duales électrofluorescentes, analogues des neurotransmetteurs endogènes (dopamine, sérotonine…) appelées Faux Neurotransmetteurs Fluorescents (FNF) ont été développées et synthétisées. Grâce à l’utilisation de ces molécules, il semble envisageable de mettre en évidence de manière unique certains aspects physico-chimiques du phénomène de libération cellulaire, afin de permettre une meilleure compréhension de ce processus biologique.
Références
[1] Chernomordik L., Chanturiya A., Green J., Zimmerberg J., The hemifusion intermediate and its conversion to complete fusion: regulation by membrane composition, Biophysical Journal, 1995, 69, p. 922-929 ;Amatore C., Arbault S., Bouret Y., Guille M., Lemaître F., Verchier Y., Regulation of Exocytosis in Chromaffin Cells by Trans-Insertion of Lysophosphatidylcholine and Arachidonic Acid into the Outer Leaflet of the Cell Membrane, ChemBioChem, 2006, 7, p. 1998-2003.
[2] Kisler K., Kim B., Liu X., Berberian K., Fang Q., Mathai C., Gangopadhyay S., Gillis K., Lindau M., Transparent Electrode Materials for Simultaneous Amperometric Detection of Exocytosis and Fluorescence Microscopy, Journal of Biomaterials and Nanobiotechnology, 2012, 3, p. 243-253 ; Meunier A. Jouannot O., Fulcrand R., Fanget I., Bretou M., Karatekin E., Arbault S., Guille M., Darchen F., Lemaître F., Amatore C., Coupling amperometry and total internal reflection fluorescence microscopy at ITO surfaces for monitoring exocytosis of single vesicles, Angew. Chem. Int. Ed., 2011, 50, 5081-5084.
[3] Gubernator N G et al., Fluorescent False NeurotransmittersVisualize Dopamine Release fromIndividual Presynaptic Terminals, Science, 2009, 324, p. 1441-1444 ;Lee M., Gubernator NG., Sames. D., J. Am. Chem. Soc., 2010, 132, p.8828-8830.
[4] Liu X., Savy A., Maurin S., Grimaud L., Darchen F., Quinton D., Labbé E., Buriez O., Delacotte J., Lemaître F., Guille-Collignon M., A dual functional electroactive and fluorescent probe for coupled measurements of vesicular exocytosis with high spatial and temporal resolution, Angew. Chem. Int. Ed., 2017, 56, p. 1–6 ;Liu X., Hu L., Pan N., Grimaud L., Labbé E., Buriez O., Delacotte J., Lemaître F., Guille-Collignon M., Coupling Electrochemistry and TIRF-Microscopy with the Fluorescent False Neurotransmitter FFN102 Supports the Fluorescence Signals During Single Vesicle Exocytosis Detection, Biophysical Chemistry, 2018, 235, p. 48-55.
[5] Lemaître F., Guille-Collignon M., Applications biologiques du couplage de la microscopie de fluorescence et de l’électrochimie, Actualité Chimique, 2015, p. 400-401.